
植物修复相对于机械修复而言是一种更安全、更好的修复方法,但植物作为修复方法有其局限性。植物作为活的有机体,其在组织中积累污染物的数量是有限的。植物和人类一样,也不是万能的。植物修复的另一个缺点是,与物理化学过程相比,它通常是一个较慢的方法,可能需要被视为一个长期的修复过程(Cunningham, Berti & Huang, 1995)。天然的超富集植物通常只积累特定的污染物。大多数植物生长缓慢,生物量小(Cunningham et al. 1993)。特别是金属超富集是一种相对少见的现象,只发生在少数陆生植物中。目前已鉴定出约400种天然超富集植物,占被子植物总数的不到0.2% (Baker et al. 2000)。
植物修复相对于机械修复而言是一种更安全、更好的修复方法,但植物作为修复方法有其局限性。植物作为活的有机体,其在组织中积累污染物的数量是有限的。植物和人类一样,也不是万能的。植物修复的另一个缺点是,与物理化学过程相比,它通常是一个较慢的方法,可能需要被视为一个长期的修复过程(Cunningham, Berti & Huang, 1995)。天然的超富集植物通常只积累特定的污染物。大多数植物生长缓慢,生物量小(Cunningham et al. 1993)。特别是金属超富集是一种相对少见的现象,只发生在少数陆生植物中。目前已鉴定出约400种天然超富集植物,占被子植物总数的不到0.2% (Baker et al. 2000)。
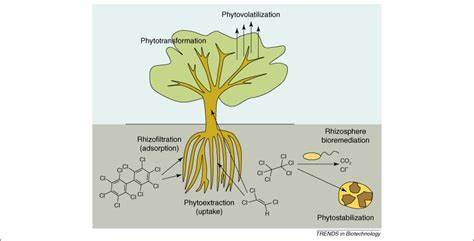
由于天然超富集体的明显局限性,如罕见、生长缓慢和富集能力有限,变基因植物的使用变得越来越重要。由于亲本之间的性不亲和性,传统的育种方法被认为是不可行的自然增加金属的提取(Lasat, 2002)。通过生物技术,已经培育出能够大量积累高浓度有毒金属并将其从根转移到芽或将其降解的植物。例如,Rugh等人(1998)对含有两种细菌基因mer A和mer B的黄杨进行了改造,以从污染土壤中富集汞并对其解毒(Datta & Sarkar, 2003)。mer B在变基因杨树的功能是催化甲基汞释放汞离子,然后由mer A转换成单质汞,比汞离子毒性小、易挥发的单质汞被释放到大气中(Datta & Sarkar, 2003)。Pilon-Smits等(1999)在印度芥菜中过度表达APT-硫酸酶(APS)基因,变基因植株的APS活性是野生型的4倍,积累的硒量是野生型的3倍。Dhankar等人(2002)报道了一种基于基因的策略,用于修复污染土壤中的砷,其中两种细菌基因在拟南芥中过度表达。一种是编码砷酸盐还原酶的大肠杆菌arsC基因,该基因将砷酸盐还原为亚砷酸盐,与光诱导的大豆rubisco启动子耦合。第二种是大肠杆菌-ECS连接到一个强组成的肌动蛋白启动子。变基因植株表现出更强的砷耐受性。在砷污染的土壤中生长时,这些植物的鲜重累积量是野生型植物的4-17倍,积累量为2-3倍。
然而,变基因植物在植物修复中的广泛使用经常遇到来自环保倡导组织的阻力,如绿色和平组织,他们声称变基因作物的环境影响尚未得到最终评估。变基因广泛使用缓慢的另一个原因是,许多国家仍然没有关于开发和使用这种技术的立法指导方针。还有人担心,变基因植物的花粉可能会污染非目标物种,造成未知的后果。在变基因植物中插入终止基因可以降低变基因植物对非目标植物的污染风险。携带终结基因的植物和其他植物一样生长,但有一个关键的例外,它们产生的种子不能长成任何植物。
(内容来源于网络,如有侵权请联系删除)
相关推荐:
边坡生态修复植物试种项目设计方案
知识点:变基因植物在植物修复中的应用