
研究背景 膜分离技术利用具有选择透过性的半透膜实现多组分体系的分离和纯化,在生物、医药、石油化工、食品、环保等领域都有广泛应用。 近年来膜分离在污废水处理、海水淡化、饮用水净化、纯水制备等水处理应用中已成为重要技术。膜污染是水处理膜分离应用面临的主要问题之一。处理物料中的微粒、胶体、溶解性有机物、金属沉淀物、微生物等污染物造成表面沉积和膜孔堵塞,使膜的透水性和分离特性发生不可逆变化,将降低产水量和出水水质,提高跨膜压差(transmembrane pressure,TMP),并缩短膜的寿命。
研究背景
膜分离技术利用具有选择透过性的半透膜实现多组分体系的分离和纯化,在生物、医药、石油化工、食品、环保等领域都有广泛应用。 近年来膜分离在污废水处理、海水淡化、饮用水净化、纯水制备等水处理应用中已成为重要技术。膜污染是水处理膜分离应用面临的主要问题之一。处理物料中的微粒、胶体、溶解性有机物、金属沉淀物、微生物等污染物造成表面沉积和膜孔堵塞,使膜的透水性和分离特性发生不可逆变化,将降低产水量和出水水质,提高跨膜压差(transmembrane pressure,TMP),并缩短膜的寿命。 根据污染物的特性,膜污染分为无机污染、有机污染和生物污染等类型。 其中,微生物可在膜上附着生长,形成的生物膜及其蛋白质、多糖等代谢产物还会改变膜表面特性,加速其他类型的污染过程。因此, 生物污染比有机污染和无机污染影响更为严重。 膜生物污染控制对膜技术在水处理应用中的发展具有重要意义。
膜污染控制主要通过膜材料选择、组件结构设计、料液参数、操作条件和膜清洗再生等方法实现。 基于生物方法的膜材料改性和膜污染控制是膜分离领域研究的前沿方向。细菌可通过产生自诱导物(autoinducer,AI)调控细胞密度依赖型的基因表达,从而调节细菌群体的生理特征,如产生胞外聚合物(extracellular polymeric substances,EPS)、形成生物膜等,这一现象称为群体感应(quorum sensing,QS)。通过干扰和阻断细菌的QS通路,可抑制相关基因的表达,该群体淬灭机制能抑制生物膜的形成,其在膜污染控制中的研究已引起关注。本文对 近年来基于群体感应的膜生物污染及膜材料改性等研究 进行综述,为水处理应用中的膜生物污染控制和抗污染膜材料的开发提供新思路。
摘 要
生物污染是水处理膜分离应用面临的主要问题之一。生物膜的形成受细菌群体感应系统调控,群体感应抑制是控制膜生物污染的新兴技术。介绍了 群体感应机制及其参与生物膜形成的有关研究。 通过干扰和阻断细菌的信息交流通路,可阻止群体感应依赖型基因表达从而抑制细菌的特定群体行为。综述了 基于细菌群体感应和群体淬灭的水处理膜生物污染控制研究。 考察了 各类群体感应抑制剂在膜法水处理系统中的应用,以及抑制剂固定化、膜材料改性等 研究进展。展望了 群体感应理论在膜生物污染控制中的研究方向 。
01
群体感应机理
群体感应是细菌之间进行信息交流从而调节群体行为的一种方式。Nealson等对海洋费氏弧菌( Vibrio fischeri )生物发光的研究发现荧光素酶在细胞增长到较高密度后爆发性合成。之后Fuqua等提出群体感应的概念,指细菌能产生、释放和检测特定化学信号分子(即AI),以此感知周围环境的细胞密度,当细菌密度达到一定水平,信号分子浓度积累到一定阈值,就会启动特定基因的表达,表现与单个细胞不同的群体生理行为,如生物发光、毒素分泌、孢子产生和生物膜形成等。
根据信号分子和感应机制的不同,QS系统主要包括以下几类(如图1)。
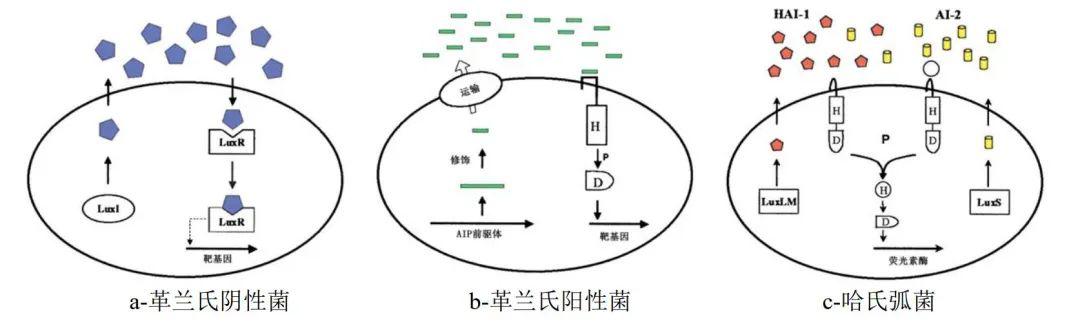
图1 细菌的群体感应系统
1)革兰氏阴性菌的LuxI/LuxR型QS系统,以酰基高丝氨酸内酯(acyl homoserine lactone,AHL)为信号分子。 细胞由LuxI型蛋白合成AHL,分泌到胞外的AHL随细胞密度上升积累到阈值,扩散进入胞内与LuxR型受体蛋白结合,形成的LuxR-AHL复合体结合靶基因的启动子,激活相关基因转录。AHL分子由一个高丝氨酸内酯环和不同分子量、不同结构的酰基侧链组成,LuxR蛋白对其同源AHL有较强结合特异性,从而实现细胞种内通讯。自 V. fischeri 中发现自诱导物AHL以来,AHL介导的群体感应也在铜绿假单胞菌( Pseudomonas aeruginosa )等革兰氏阴性菌中大量发现并开展深入研究。
2)革兰氏阳性菌的双组分QS系统,以经过修饰的寡肽类分子(autoinducing peptide,AIP)为信号分子 。AIP前驱体在胞内经修饰后形成稳定、特异的AIP,由ATP结合转运组件或其他膜通道蛋白运输到胞外,胞外AIP浓度到达阈值,信号分子被细胞膜上的激酶识别,激酶的组氨酸残基发生磷酸化,磷酸基团由胞内的天冬氨酸残基转运至受体蛋白,蛋白磷酸化后结合靶基因并触发基因表达。感应体系由于细胞膜激酶对AIP的高度选择性而同样具有特异性。
3)细菌种间交流的QS系统。 AHL和AIP参与细菌的种内群体感应,称为AI-1。而由AI-2介导的细菌种间交流在革兰氏阴性和阳性菌中均有发现。在较为典型的革兰氏阴性菌哈氏弧菌( Vibrio harveyi )生物发光QS通路中,与AHL类似的HAI-1由LuxLM合成并由LuxN感应;呋喃酰硼酸二酯类化合物作为AI-2,由LuxS合成并由LuxP和LuxQ感应,其识别方式与革兰氏阳性菌的双组分激酶识别系统相同,LuxN和LuxQ通过磷酸转移酶LuxU将信号传递至调节蛋白LuxO,在LuxR协助下启动基因表达。
群体感应已在铜绿假单胞菌、金黄色葡萄球菌、大肠杆菌、沙门氏菌、枯草芽孢杆菌和豆科根瘤菌等很多细菌中发现。 细菌使用多种信号分子和受体蛋白进行多个回路的并行或串联作用,体现出复杂的QS机制。许多革兰氏阴性菌使用AHL的同时产生AI-2。同样,许多革兰氏阳性菌具有AIP和AI-2通路。细菌通过合成和感应多种信号分子评估种内密度和其他种群的密度,不断调节群体行为。
02
群体感应对生物膜形成的作用
生物膜的形成是QS研究中最受关注的生理特性之一。生物膜是附着在载体表面由EPS包裹、有组织生长的细菌聚集体。 生物膜的形成包括可逆粘附定殖、不可逆粘附集聚、细菌繁殖、生物膜成熟及生物膜老化脱落等阶段。 细菌通过群体感应调控生物膜形成及菌群在生物膜内的行为。 Davies等在 P.aeruginosa 研究中发现信号分子参与生物膜的形成过程,其信号突变株( lasI 突变株)形成的生物膜呈扁平状,对杀菌剂十二烷基硫酸钠相比野生型更敏感;在投加外源信号分子后,突变株生物膜呈正常状态;研究指出QS系统参与了生物膜的分化过程。之后Bassler等发现 P.aeruginosa 有两个典型的LuxI/LuxR系统(LasI/R 和RhlI/R),分别产生和检测3OC12-HSL和C4-HSL。RhlR同时指导RhlI依赖型和非RhlI依赖型调节子,在缺乏RhlI时,RhlR控制生物膜形成所需基因的表达。
群体感应对生物膜各阶段调控作用的研究已有报道。 金黄色葡萄球菌通过副基因调节器QS系统协调表面粘附行为。群体感应通过控制EPS合成基因来调控霍乱弧菌( Vibrio cholerae )生物膜的成熟,相比野生型,Δ hapR 突变株中 vpsA-K 和 vpsL-Q EPS操纵子的表达被诱导,生物膜被强化,而Δ luxO 突变株中的操纵子被下调,形成的生物膜受损。液化沙雷氏菌( Serratia liquefaciens )的集群细胞分化受N-丁酰基-L-高丝氨酸内酯和N-乙基-L-高丝氨酸内酯调控,其AHL合成基因 swrI 修饰后突变株不表现群集运动,形成的生物膜稀薄,但添加外源AHL后恢复群集运动。黄单胞杆菌( Xanthomonas campestris )的胞外甘露糖苷酶参与黄原胶的裂解和聚集体的溶解扩散,其合成受DSF/ rpf QS系统调节。 V.cholerae 的QS系统在高细胞密度时会抑制生物膜的形成,促使生物膜在不良条件时解体。
03
基于群体感应的膜污染控制
1. 群体感应的抑制
群体淬灭是通过干扰细胞间的信息交流,阻止QS依赖型基因表达而抑制细菌的特定群体行为。根据QS系统的组成和特点,群体感应抑制有以下几种途径。 1)抑制信号分子的产生。 AHL在胞内通过酰基-酰基载体蛋白和S-腺苷甲硫氨酸结合合成,参与合成的酶包括AHL合成酶、烯酰基载体蛋白还原酶、酰基载体蛋白酶等。AIP通过前体肽的断链和修饰合成。通过消除底物或抑制合成酶活性可阻断信号分子合成。例如,S-腺苷甲硫半胱氨酸同系物对 P.aeruginosa 信号合成酶RhlI活性有抑制效果,可抑制信号分子合成。 2)降解信号分子。 具有信号分子降解活性的细菌、酶和化合物已有报道。You等从海洋放线菌中分离出多种菌株,能降低AHL活性,抑制 V.harveyi 、创伤弧菌( Vibrio vulnificus )和鳗弧菌( Vibrio anguillarum )生物膜形成。Augustine等从芽胞杆菌( Bacillus spp.)中获得的AHL内酯酶(AiiA)可抑制 V.cholerae 生物膜形成。Lin等从罗尔斯通氏菌( Ralstonia sp. XJ12B)中合成的酰化酶AiiD可降解长链3OC12-HSL和短链C4-HSL。HClO、HBrO等化合物也可通过卤代反应降解3-oxo-AHL。 3)阻止信号分子与受体蛋白的结合。 通过降低受体蛋白活性或引入信号分子类似物竞争结合受体蛋白可以抑制细菌QS系统。卤代呋喃酮能使受体蛋白失活而抑制 P.aeruginosa 生物膜形成和毒素产生。黄芩苷和大黄素与信号分子有相似结构,可竞争结合受体蛋白TraR,抑制 P.aeruginosa 生物膜形成,其结合体能改变蛋白构象使之易于水解,进而阻断由QS控制的反应及功能表达。
2. 基于群体感应的水处理膜污染控制
细菌在水处理膜表面滋生形成生物膜,影响系统的高效稳定运行。采用群体淬灭方法可抑制生物膜形成,并减少化学杀菌剂引起的细菌抗性等问题。Xu等在厌氧膜生物反应器(membrane bioreactor,MBR)中发现游离菌团中低丰度的AHL介导的 Rhodocyclaceae;g- 在生物膜形成初期易于造成污染,而群体淬灭可延缓生物膜的初始形成,并改变成熟生物膜的菌种结构。目前已有研究将QS理论应用于膜污染控制中,并开发多种群体感应抑制剂,以期获得更高效安全的生物污染控制方法。
1) 利用群体感应抑制化合物控制膜污染
人工合成和天然的QS抑制剂对膜生物污染的控制 研究已有报道。呋喃酮类化合物可通过钝化las和rhl受体蛋白抑制P. aeruginosa等细菌的生物膜形成。经过修饰的内酯类、邻苯三酚、噻唑烷二酮类化合物等也可通过与载体蛋白竞争结合抑制群体感应。许多生物中含有群体淬灭功能的化合物,其中一些具有信号分子类似结构的化合物能够竞争结合受体蛋白;部分天然化合物能够通过降解LuxR/LasR受体蛋白阻断QS通路。Ponnusamy等研究显示香草醛对C4-HSL、3-oxo-C8-HSL、C6-HSL、C8-HSL、C14-HSL和C10-HSL均有抑制活性,从反渗透(reverse osmosis,RO)膜生物膜中分离的嗜水气单胞菌( Aeromonas hydrophila )在添加香草醛的培养基中生长受限,香草醛浓度为0.25 mg/mL时可减少46.3%的生物膜量形成。Katebian等通过表面沉积将香草醛和肉桂醛负载于RO膜SW30XLE和SWC5表面,在接种 Alteromonas sp .和 Shewanella sp. 的合成海水过滤中,通量下降分别从50%减至34%(SW30XLE)和从22%减至15%(SWC5),改性后膜表面的多糖产生量、活菌和死菌分别减少15%、58%和61%。Xu等研究表明D-酪氨酸可抑制活性污泥微生物AI-2、eDNA、多糖和蛋白质的合成,从而减少在亲水玻片和疏水聚丙烯载体表面的附着,投加6 mg/L的D-酪氨酸可减少生物附着量达22%,底物分析显示D-酪氨酸对底物去除无明显影响。Siddiqui等将萎叶提取物加入MBR处理合成印染废水,相比对照组以及C6-HSL强化的MBR,TMP上升速度减缓,且膜表面生物膜内检测到的AI-2浓度减少约40%。
具有群体淬灭特性的化合物在膜污染控制中已取得成效,化合物抑制剂具有较稳定的性质,可投加至反应器或通过常规方法用于膜改性,实际应用可行性高。 已报道的化合物大部分在干扰生物膜形成时对细菌的生长无抑制作用,不产生抗性,同时保持细菌对底物的降解效率。Kappachery等将RO膜在投加0.18 mg/mL香草醛的CDC生物膜反应器中培养,3天后较无香草醛时的 A. hydrophila 生物膜覆盖度、平均厚度、总生物量和总蛋白质分别降低93%、97%、96%和97%;但表面预形成生物膜的RO膜在香草醛反应器中培养1天后生物膜无明显变化。水处理应用中,活性污泥和生物膜中的EPS对抑制剂有抵御作用,可减少抑制剂对其功能菌活性的破坏,这将有利于群体淬灭在生物膜-膜生物反应器等复合工艺中的应用。除香草醛、肉桂醛等之外,玫瑰茶多酚、香芹酚等天然QS抑制剂对生物膜形成也有抑制效果,有利于提高群体淬灭在水处理应用中的安全性。但部分化合物的作用是通过与相似分子结构的信号分子竞争结合受体蛋白,淬灭效果受限于信号分子侧链结构和长度等因素。因此该类抑制剂对不同菌种的QS通路干扰作用也有差异,对混合菌种培养体系的影响有待进一步考察,为其在实际膜法水处理中的应用提供指导。
2) 利用群体淬灭酶控制膜污染
群体淬灭酶可降解QS信号分子、干扰信号分子的合成或钝化受体蛋白,从而抑制QS通路。 已有研究的淬灭酶包括AHL内酯酶、酰基转移酶和氧化还原酶等,它们作用于AHL分子内酯环或侧链等多个位点,使内酯环开环生成酰化高丝氨酸,或使酰基侧链断裂转化成高丝氨酸内酯环和脂肪酸,或催化还原酰基侧链形成羟基高丝氨酸,从而降解或灭活信号分子。
酰基转移酶等群体淬灭酶在膜生物污染控制中的研究开展较早且有效性已得到验证。 Yeon等观察到MBR膜表面生物膜中AHL浓度随运行时间增加,并与TMP上升正相关;将10 mg/L酰基转移酶加入MBR后生物污染引起的TMP升高减缓,形成的生物膜中AHL浓度减少(图2)。 考虑到长期运行下游离酶的活性和流失,已有研究采用载体固定淬灭酶以提高处理效果稳定性。 Yeon等采用磁性二乙烯基苯-甲基丙烯酸缩水甘油酯离子交换树脂载体固定酰基转移酶(图3),控制MBR的生物膜集聚;投加酶载体的MBR运行48 h后TMP保持在10 kPa左右的初始值,未投加酶载体MBR的TMP增加到30 kPa左右。Lee等在带磁性纳米颗粒的球状介孔二氧化硅载体中固定酰基转移酶,在低酶量和高有机负荷的条件下也能保持淬灭酶的抗污染活性,将载体与微滤膜混合培养,膜表面 P.aeruginosa 生物膜的形成减缓。Jiang等将固定酰基转移酶的海藻酸钠胶囊投入MBR,使TMP增长速度从0.611 kPa/h降至0.075 kPa/h,膜表面EPS累积量减少50%,而COD、氨氮、总氮等污染物去除率无明显影响,同时可提高污泥沉降性能。使用QS抑制剂进行膜改性也是控制膜污染的有效方法之一。Kim等通过合成壳聚糖-酰基转移酶基质,将酰基酶固定在纳滤膜表面,经过38 h的操作,改性纳滤膜的通量仍保持在初始通量的90%以上,而未改性的膜在12 h后通量开始下降,并持续下降至60%;改性膜表面生物积累量从1.15 μm3/cm2减少至0.06 μm3/cm2(图4)。Kim等通过酶吸附沉淀交联法将酰基转移酶固定于碳纳米管上,然后采用聚多巴胺将其负载于PVDF膜上,改性后膜表面生物膜形成得到抑制,在过滤接种P. aeruginosa的料液时TMP增长至25 kPa所需时间从11 h延长至18 h。Zhu等制备了酰基转移酶/氧化石墨烯改性PVDF膜,改性后膜表面生物积累量减少84%,有效抑制了生物膜的形成。
图2 投加酰基转移酶和AHL的MBR测试结果
图3 层层自组装制备离子交换树脂磁性酶载体示意
图4 膜表面 P.aeruginosa 生物膜的激光共聚焦扫描显微镜
相比化合物抑制剂, 群体淬灭酶可降低反应所需活化能,大幅提高淬灭效率。 酶可作用于范围更广的同类信号分子合成、累积和受体蛋白结合,有利于淬灭混合菌群的QS通路。但通过群体淬灭降低生物污染是可逆的,群体淬灭停止后膜分离性能将不会被改善,因而需要不断引入抑制剂实现长效的膜污染控制,使用淬灭酶的运行成本较高。另外, 酶对高温、酸碱等环境条件极为敏感,利用载体固定化或结合膜材料改性有利于维持酶的活性,需进一步研究固定方法和载体结构的优化,促进酶的控制释放和活性保持。
3)利用群体淬灭菌控制膜污染
群体淬灭菌的应用也是膜污染控制领域的研究方向之一,可克服群体淬灭酶在活性和高成本等方面的问题。 群体淬灭菌能以多种信号分子为碳源和能源,如红平红球菌( Rhodococcus erythropolis )可降解酰基侧链长C8-C14的3-oxo-AHL等信号分子。已有研究从活性污泥中分离出多种群体淬灭菌。Oh等从污水处理MBR中分离出的红球菌( Rhodococcus sp. BH4)对C8-HSL具有分解作用,可使膜表面生物膜更稀薄。Cheong等从污水处理厂活性污泥中纯化得到可产生酰基转移酶的假单胞菌( Pseudomonas sp. 1A1),对C10-HSL、C12-HSL等长链AHL和3-oxo-C12-HSL降解能力更高。Kampouris等从市政污水处理厂活性污泥中筛选得到乳酸菌( Lactobacillus sp. SBR04MA),该菌株使50 μM C6-HSL在9 h内完全降解。Ham等从MBR中分离的肠球菌( Enterococcus sp. HEMM-1)能分泌内酯酶降解AHL,在CDC生物膜反应器中可使微滤膜表面活性污泥形成的生物膜厚度从25.98 μm减至13.41 μm。
群体淬灭菌在膜污染控制中的实际应用也在探索中。Oh等将分泌AHL内酯酶的重组大肠杆菌和群体淬灭菌BH4分别注入聚乙烯中空纤维膜(图5a),将该微生物容器置于MBR后能减缓TMP上升,其中BH4的MBR运行40天后TMP为20 kPa,无BH4的MBR为50 kPa。经Oh等分析,BH4通过分泌AHL内酯酶使C8-HSL开环而被降解,BH4对考察的8种AHL在10 min内降解率为10-80%,其中酰基侧链含oxo基团的AHL降解率较低,侧链越长越易被降解;信号分子在BH4胞内降解较胞外明显更高。Weerasekara等将BH4微生物容器置于MBR中,联合使用淬灭菌和氯清洗的反应器50天后TMP升至20 kPa,只使用氯清洗的反应器则升至50 kPa;该联合方法使过滤能耗减少约74%;且Cl2当量浓度100 mg/L的次氯酸钠对BH4的淬灭活性无影响。Kim等将BH4包埋于可自由移动的海藻酸钠微球中(图5b),通过物理摩擦和群体淬灭的双重作用减轻膜表面微生物附着,使TMP升至70 kPa的时间延长10倍。为减少海藻酸钠微球在水中的分解,Lee等制备了BH4的海藻酸钠/聚乙烯醇微球,应用于中试规模的一段式MBR和三段式MBR中,实际污水处理结果显示,投放淬灭菌微球后一段式和三段式MBR的TMP从10 kPa上升至20 kPa的时间分别从~15天延至~28天、从18天延至35天;一段式MBR淬灭菌的C8-HSL降解活性在前25天减少40%但60天后逐渐恢复至初始值,三段式MBR淬灭菌活性在100天内降低10-20%;淬灭菌微球不影响出水水质,还可减少膜清洗的曝气能耗。Lee等将BH4制备成海藻酸钠/聚乙烯醇中空圆柱,以提高载体内的传质效率(图5c)。Kampouris等将乳酸菌SBR04MA制备成海藻酸钠微球,在MBR中可减少膜表面溶解性微生物产物和EPS的产生,使临界通量从8.3 L/m2 h提高到24.25 L/m2 h,且不影响COD去除效率。Oh等也试验了具有淬灭功能的重组大肠杆菌对RO膜污染的抑制,无淬灭菌的RO系统TMP在115 h时增加一倍,其中前97 h增加20%而后18 h增加80%,接种淬灭菌的系统115 h时TMP增加33%,但136 h时为初始TMP的两倍。除向系统中投加游离态菌株以外,Shah等将混合BH4的海藻酸钠/聚乙烯醇溶液涂覆在聚砜和PVDF中空纤维膜上,改性后膜初始水通量降低了34-47%,但膜污染被延缓了57-67%,将是提高膜材料抗污染性能的新途径。
图5 群体淬灭菌固定化载体
群体淬灭菌在水处理膜污染控制中具有很大的应用潜力。淬灭菌可分泌多种淬灭酶,以不同种类的信号分子为生长底物,降解信号分子,相比其他抑制剂更有利于抑制混合菌群引起的膜污染。虽然淬灭菌活性弱于直接添加淬灭酶,但成本低且对操作条件适应性更强,有利于提高实际应用可行性。 群体淬灭菌在污废水处理、海水淡化等系统中广泛存在,从水处理系统中筛选高效的淬灭菌将是研究热点之一。 另外, 可采用生物刺激法原位培养群体淬灭菌。 已有研究将与γ-已内酯加入MBR中以激活淬灭菌,可使系统中AHL内酯酶生成基因增多,使膜污染得到控制。
04
结束语
膜生物污染是膜技术在水处理应用中亟待解决的关键问题。通过干扰细菌群体感应系统抑制生物膜的形成已被证明是有效控制生物污染的新技术,在膜污染控制中具有良好的应用前景。现有研究对群体感应系统机制和对生物污染的调控方法仍较局限。今后需对水处理膜分离中的群体淬灭机理和应用开展深入探讨,研究方向包括以下方面:
1)针对水处理应用,研发有效、稳定的群体感应抑制化合物和群体淬灭酶,从水处理系统中筛选或采用基因重组方法构建新型群体淬灭菌。 目前研究以AHL介导的系统为主,用于其他种内和种间QS通路的抑制剂较少。鉴于进水中的微生物多样性,需探索抑制剂对优势菌和混合菌群的作用,评价膜污染缓解效果,并考察其对出水水质和环境安全的影响。
2)研究群体感应抑制剂的引入方法。 固定化抑制剂在污染控制效果和稳定性方面较游离态更有优势。对于载体固定化抑制剂,由于淬灭效率受限于反应器到载体内部的传质速率,需研究新型的载体制备技术和载体结构。通过生物刺激等方法可原位强化水处理系统的群体淬灭菌。另外,采用QS抑制剂进行膜材料制备和改性,改善抑制剂释放的可控性,将有利于精准地控制膜表面的微生物集聚,提高膜抗污染能力和清洗效率。
3)生物膜法和活性污泥等生物处理是水处理中的主要单元之一。 细菌的集聚对菌胶团生长、载体挂膜、污泥絮凝沉降等过程有重要作用。例如,在生物膜-膜生物反应器等生物膜法和膜分离的结合工艺中,细菌需有较强的EPS分泌和生物膜形成能力使其在载体表面附着。好氧颗粒污泥法中,群体感应在颗粒形成初期起诱导作用,可促进微生物附着生长并形成颗粒。研究表明群体感应抑制剂对MBR的污染物去除效率无明显影响。由于工程应用中膜分离会与活性污泥和生物膜等多种生物处理单元联用,今后应结合实际应用,研究群体感应抑制剂对其他工艺的水处理效果的作用,降低抑制剂对生物处理单元的不利影响,并减少抑制剂投加量和应用成本。